
30 de diciembre de 2010
24 de diciembre de 2010

18 de diciembre de 2010
Pío del Río Hortega (1882-1945)
Pío del Río Hortega es, probablemente después de Santiago Ramón y Cajal (1852-1934), la personalidad más destacada de la neurohistología española de todas las épocas.
Nació en Portillo (Valladolid) y en la Facultad de Medicina vallisoletana se licenció en 1905 y, más tarde se doctoró.
Aunque eventualmente ejerció la medicina (fue médico de su pueblo natal), ese trabajo le resultó tan negativo que optó por realizar el doctorado como una especie de “respiro a pulmón lleno”. Su gran interés científico lo constituyó la investigación en el campo de la histología y así, aunque realizó trabajos sobre la histología patológica de las tumoraciones nerviosas, su discípulo Ortiz Picón lo definió más como histólogo que como anatomopatólogo.
En 1913 se traslada a Madrid y trabaja en el Laboratorio de Histopatología del Sistema Nervioso que dirigía Nicolás Achúcarro Lund (1880-1918). Una vez en la capital de España, obtiene una beca para ampliar conocimientos en París y Berlín, pero la Primera Guerra Mundial le obliga a volver a Madrid. Tras el fallecimiento de Achúcarro se hace cargo de la dirección del laboratorio y, en poco tiempo, don Pío hace de él un centro de investigación de gran categoría internacional. Prueba de ello es que grandes científicos se trasladaron a Madrid para trabajar en su laboratorio; es el caso, por ejemplo, del norteamericano Wilder Penfield.
Al iniciarse la Guerra Civil se exilia a Francia donde trabaja como histopatólogo en el Hospital de la Pieté de París; posteriormente marcha a Gran Bretaña para investigar en la Universidad de Oxford (de la que fue nombrado doctor honoris causa). Finalmente, en 1940, como consecuencia de la Segunda Guerra Mundial, va a Buenos Aires y dirige, hasta su muerte en 1945, un laboratorio histológico dependiente de la Institución Cultural Española de la capital argentina.
En el mundo de la ciencia don Pío destacó por la precisión de sus tinciones tisulares. En efecto, fue alterando la tinción de Achúcarro (que utilizaba tanino y plata amoniacal) de tal manera que consiguió realizar importantes estudios de la estructura anatómica de las neuronas y de la neuroglía, desarrollando su famoso método del carbonato de plata amoniacal (1918). Estos métodos de tinción permitieron el descubrimiento de unas células que Ramón y Cajal había llamado tercer elemento o glía adendrítica; los otros elementos eran las neuronas (primer elemento) y la neuroglía (segundo elemento), donde se encontraban formas celulares como los astrocitos de la sustancia gris y los de la sustancia blanca. En efecto, Río Hortega demostró la existencia de dos estirpes celulares diferentes en este tercer elemento: las células de oligodendroglía y las células de microglía.
Además, en la década de los 20, el investigador castellano publicó numerosas investigaciones sobre la morfología y génesis de estas células que le valieron renombre internacional. Así, dos importantes científicos alemanes (Metz y Spatz) ya hablaban en esos años de las “células de Hortega” para designar a la microglía.
7 de diciembre de 2010
El dilema del sueño
Todos los animales vertebrados duermen. Aunque los anfibios y peces no lo hacen con las características que definen el sueño en los demás animales, también manifiestan periodos de inactividad semejantes al sueño.
Y es que el sueño debe ser necesario ya que algunas especies de mamíferos probablemente estarían mejor en vigilia permanente que soportando los peligros que tienen cuando duermen. Es el caso, por ejemplo, del delfín de Índico, que duerme hasta 7 horas al día a pesar de ser ciego y de nadar constantemente en las oscuras aguas del golfo Índico. Su adaptación al sueño es muy peculiar ya que duerme dando pequeñas “cabezadas” de menos de un minuto.
Otros animales presentan estrategias de sueño diferentes. Así, por ejemplo, el delfín de morro de botella posee dos hemisferios cerebrales que duermen por turnos: el derecho unas veces y el izquierdo otras. De esta forma un hemisferio cerebral siempre está vigilante.
La conclusión consecuente de estos y otros hechos es que si esta conducta, la del sueño, fuera innecesaria muchos animales carecerían de ella ya que, evolutivamente, habría desaparecido en buena parte de ellos. No obstante, todos los vertebrados duermen.
Sabemos que, en nuestra especie, la privación del sueño nos provoca alteraciones en la percepción y, cuando la ausencia del sueño es demasiado larga, algunas personas llegan a tener, en ocasiones, alucinaciones ligeras y problemas para realizar tareas que necesitan concentración. Sin embargo numerosos estudios científicos no han demostrado que el sueño sea necesario para el funcionamiento del organismo, ya que en los caso de privación prolongada no se han manifestado señales de estrés fisiológico aunque sí de percepción, problemas de concentración y alguna alucinación. Finalmente, cuando a la persona que se le ha privado del sueño se le permite dormir, duerme más en la primera noche pero, en ningún caso, recupera el sueño perdido.
Es por esto que la ciencia se encuentra actualmente con un dilema difícil: el sueño parece necesario (todos los vertebrados duermen) y, a la vez, no parece imprescindible.
25 de noviembre de 2010
Autismo y genética
Alrededor de cuatro de cada diez mil niños nacen con autismo y aunque sabemos de diversos factores ambientales que aumentan el riesgo de padecer autismo, bien es cierto que ninguno de ellos es suficiente por si mismo para que se manifieste la patología. La mayor parte de ellos está relacionado con la exposición del embrión o del feto a los efectos nocivos de algunos virus como el de la rubéola, o a sustancias químicas como el ácido valproico o el etanol.
Desde hace tiempo se sabía que el autismo tenía un componente genético. Es curioso que algunas patologías genéticas incrementan la posibilidad de padecer autismo; es el caso, por ejemplo, de la fenilcetonuria y del síndrome del cromosoma X-frágil, trastornos ambos que se expresan con bajos niveles de inteligencia. En este último caso hay que tener en cuenta que el 15% de los afectados con el síndrome del X-frágil sufre autismo.
Aunque hay un 95% de casos en los que un autista no tiene parientes con el trastorno, en su familia es más probable que se dé un porcentaje mayor de alteraciones afectivas, sociales, comportamientos estereotipados, tics, déficit de comunicación, anomalías del lenguaje, epilepsia, etc., irregularidades características de esta patología. Además, los hermanos de autistas tienen una probabilidad entre un 3 y un 8% de padecer la enfermedad, lo que contrasta considerablemente con el 0,16% de riesgo de la población no emparentada con autistas. Bien es cierto que si el autismo fuera debido a un solo alelo (dominante o recesivo) la probabilidad sería mayor. Y es que en esta enfermedad hay muchos genes implicados.
Por otra parte, para confirmar las implicaciones de los genes, hay estudios diferentes que indican que cuando una persona es autista su gemelo idéntico tiene una probabilidad entre el 60% y el 96 % de serlo también. Finalmente, hay que decir que la incidencia del autismo es 3-4 veces superior en los varones que en las mujeres.
Entre un 5 % y un 15 % de los casos de autismo presentan alteraciones cromosómicas que afectan al número o a la estructura (deleciones, duplicaciones, etc.). Las más frecuentes implican a los cromosomas 15 y al cromosoma X.
En este siglo se ha relacionado el autismo con las variantes anómalas de dos genes situados en el cromosoma X. Estos genes son responsables de la síntesis de unas proteínas que se encuentran en la superficie de las neuronas y parece que sirven para que se produzca la correcta transmisión de los impulsos nerviosos. Por otra parte, también se han descubierto variantes de genes normales localizados en los cromosomas 2, 16 y 17 que están implicados en la enfermedad.
Asimismo, en el cromosoma 7, otros investigadores han encontrado dos genes relacionados con el autismo: uno es un gen que parece que tiene una importancia fundamental en una forma de trastorno del lenguaje característica de los autistas.
Hay otro gen que se relaciona con el autismo; se denomina HOXA1 y también se localiza en el cromosoma 7. De este gen se han encontrado varios alelos diferentes que se consideran, al menos parcialmente, responsables de la enfermedad, ya que se presentan con más frecuencia en la población autista que en la que no lo es. Sin embargo, su presencia no implica la existencia de autismo y su ausencia no supone la imposibilidad de padecer el trastorno: se encuentra en un 20% de las personas normales y en un 40% de las autistas.
15 de noviembre de 2010
Endorfinas, morfinas y sistema nervioso
Durante siglos el opio ha sido punto de referencia debido a su capacidad para reducir el dolor y producir euforia. Pero desde un aspecto terminológico conviene recordar que opiáceo se refiere a todos los productos derivados de la planta emparentada con la amapola y a la que los científicos denominan Papaver somniferum, o lo que es igual: “hierba del sueño”.
El vegetal produce un fruto en cápsula, globoso, que al hacerle unos pequeños cortes suelta un líquido que cuaja en contacto con el aire, el opio, y que contiene más de
veinte alcaloides. De éstos, el más importante es la morfina, sustancia descubierta en 1804 por el farmacéutico alemán Sertürner y así llamada en honor a Morfeo, dios del sueño de la mitología griega.
La morfina fue utilizada como analgésico desde el primer momento, aunque pronto se observó un enorme problema: creaba adicción. Por tal motivo, algunos laboratorios farmacéuticos se lanzaron a la búsqueda de un principio activo similar pero que no tuviera efectos indeseables. Así las cosas, la compañía Bayer inició en 1898 la comercialización de una sustancia, que había sintetizado en sus laboratorios, la heroína. Sin embargo, la nueva molécula, además de crear adicción, ejercía un efecto más rápido que la morfina.
En el año 1975, Hughes y Kosterlitz sorprendieron a la comunidad científica al descubrir la existencia de algunas sustancias que tenían las mismas propiedades de la morfina y… que eran sintetizadas por el propio organismo; por ello se las denominó genéricamente endorfinas, y más tarde opiáceos endógenos, para diferenciarlos de los que no eran fabricados por el cuerpo, o exógenos. Después, y en muy poco tiempo, se descubrieron otras sustancias similares: encefalina, b-endorfina, dinorfina, etc.
Curiosamente todos los opiáceos endógenos se localizan en zonas del sistema nervioso que guardan alguna relación con el procesamiento o la modulación del dolor.
¿Por qué los opiáceos actúan sobre el organismo?
Los opiáceos, endógenos o exógenos, podrán estimular a todas las neuronas que tengan receptores para ellos. Pero, ¿dónde se encuentran las células nerviosas sobre las que pueden actuar la morfina o la heroína?
Estas neuronas se localizan en diferentes zonas del cerebro: el área tegmental ventral, el núcleo accumbens, el área preóptica, la formación reticular, la sustancia gris periacueductal y otras. Pues bien, las regiones que acabo de citar explican de una manera muy clara los efectos, perfectamente detectables, de los opiáceos: refuerzo, descenso de temperatura (hipotermia), analgesia, etc.
5 de noviembre de 2010
Señales externas y sistema neuroendocrino
El hipotálamo produce unas hormonas que pueden ser de dos tipos: las llamadas hormonas de liberación y hormonas de inhibición hipotalámicas. Estas sustancias se fabrican en el hipotálamo y viajan a través de vasos sanguíneos hasta la adenohipófisis, donde ejercen su efecto.
Veamos un ejemplo que explique esta “cascada” hormonal que nace en el hipotálamo. Esta estructura neuroendocrina produce una hormona que se denomina hormona liberadora de tirotropina (u hormona liberadora de TSH) que es llevada por los vasos sanguíneos hasta la adenohipófisis. Una vez allí actúa haciendo que se fabrique más TSH y que se libere esta hormona. La TSH , cuando llega a la glándula tiroidea, hace que se libere a la sangre las hormonas tiroxina y triyodotironina. Finalmente, estas hormonas tienen efectos sobre el metabolismo celular, crecimiento, etc.
Pero esta cascada puede ser alterada por factores perfectamente bien conocidos y estudiados: el frío afecta de tal manera al hipotálamo que provoca un aumento en la secreción de la hormona liberadora de tirotropina y por tanto, una mayor secreción de TSH primero y de hormonas tiroideas después. Determinados estados emocionales, como los de ansiedad, provocan un agudo descenso en la liberación de TSH y, en consecuencia, de las hormonas que produce la glándula tiroidea.

26 de octubre de 2010
Dopamina y psicosis
Aunque en neurobiología las pruebas científicas de un hecho neuronal casi nunca son concluyentes debido, principalmente, a las numerosas interconexiones entre las células nerviosas, es bastante incuestionable que la dopamina tiene una importancia fundamental en las psicosis. Lo que no es excluyente con otras teorías que postulan que, además, participan en la enfermedad otros neurotransmisores.
Así, en general, podemos afirmar que aquellos fármacos que aumentan la disponibilidad de la dopamina tienen efectos psicóticos positivos y los que disminuyen la dopamina tienen como efecto reducir los signos psicóticos positivos.
Ya desde la década de los 50 del siglo XX se sabía que la reserpina, disminuía los niveles de dopamina, y de otras monoaminas, y tenía un buen efecto antisicótico. También se conoce desde hace mucho tiempo que la cocaína y las anfetaminas aumentan las cantidades de dopamina en los espacios intercelulares de las neuronas y generan episodios semejantes a los que se presentan en personas enfermas por esquizofrenia.

La dopamina es un neurotransmisor que se acopla a los receptores correspondientes. Sin embargo, hay determinados fármacos que se unen a los receptores postsinápticos de la dopamina y, en consecuencia, no permiten la actuación del neurotransmisor. Cualquier fármaco que bloquea la neurotransmisión debida a la dopamina y evita o reduce los signos de la psicosis de denomina, genéricamente, neuroléptico.
17 de octubre de 2010
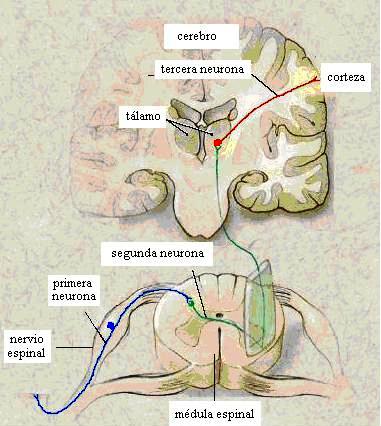
Las vías sensoriales tienen pocas neuronas
Los circuitos nerviosos que llegan a la corteza cerebral desde los receptores son vías más o menos largas, pero en ellos no suelen estar implicadas muchas neuronas. Veamos algún ejemplo.
Desde la retina ocular, donde se ubican las células receptoras visuales (conos y bastones), parten neuronas (sensitivas) que forman el nervio óptico (de cada ojo) y que finalizan en el tálamo. Aquí hacen sinapsis con otras neuronas que finalizan en la corteza visual. En resumen, un circuito visual, desde que sale del ojo, necesita dos neuronas: una que llega hasta el tálamo y otra que va desde esta región cerebral hasta la corteza cerebral.

Claro que usted pensará que esto se debe a que los receptores visuales y la corteza visual se encuentran espacialmente muy cerca. Sin embargo, aunque esto es verdad, no es por ello.
En efecto, si nos fijamos en un circuito doloroso que parte, por ejemplo, de un músculo de la pierna, los impulsos nerviosos viajan por una neurona (sensitiva) que termina en la médula espinal. Aquí conecta con otra neurona cuyo axón cruza al otro lado (decimos que decusa y, por ello, que es contralateral) y termina en el tálamo. Aquí conecta con otra neurona que finaliza en la corteza cerebral. Es un caso, por tanto, muy semejante al anterior, con tres neuronas llega la información cerebral a ser consciente.
6 de octubre de 2010
Tipos de neurotransmisores
Hay varias clases de neurotransmisores y la mayoría de ellos generan un PEP o un PIP, aunque, en algunos casos, un determinado neurotransmisor puede producir excitación o inhibición dependiendo del receptor al que se una.
a) Algunos son aminoácidos (los componentes de las proteínas).
Son los casos del aminoácido glutamato (el neurotransmisor excitatorio más abundante en nuestra especie), el aspartato, la glicina y el ácido gamma-aminobutírico (GABA); este último es el neurotransmisor inhibidor más abundante.
b) Otros son monoaminas.
En este caso hay dos grupos de neurotransmisores: las catecolaminas y las indolaminas. De entre las primeras destacamos la dopamina, adrenalina (o epinefrina) y la noradrenalina (o norepinefrina). Estos tres neurotransmisores se sintetizan a partir del aminoácido tirosina (no confundir con la hormona tiroxina). Indolamina es la serotonina, un neurotransmisor que se sintetiza a partir de otro aminoácido, el triptófano.
c) La acetilcolina es una molécula que actúa como neurotransmisor en muchas sinapsis y en las uniones neuromusculares.
d) Neuropéptidos. Numerosos péptidos (son moléculas formadas por la unión de aminoácidos. Muchos aminoácidos unidos forman una proteína.) sintetizados en las neuronas actúan como neurotransmisores. Es el caso, por ejemplo, de las endorfinas.
e) Desde hace poco sabemos que ciertos gases como el óxido nítrico y el monóxido de carbono son capaces de atravesar fácilmente las membranas y estimular la formación de un segundo mensajero.
28 de septiembre de 2010
El nuevo impulso nervioso
Los potenciales postsinápticos no responden a lo indicado en el caso de los potenciales de acción. Esto es, no se rigen por la "ley del todo o nada", sino que las amplitudes (de excitación o de inhibición) dependerán de las intensidades que las provoquen, es decir, son potenciales graduados.
Por otra parte, esta respuesta, que se inicia allí donde termine un botón terminal (sea dendrita o cuerpo celular generalmente) va decayendo progresivamente según viaja en dirección al cono axónico, la estructura cónica que se encuentra en la unión del cuerpo celular y del axón. Así, si el neurotransmisor ha generado un PIP de –75mV, según viaja y se aleja del punto inicial, esta onda de hiperpolarización irá atenuándose a –74mV, –73mV... De la misma forma, si se produce un PEP de –65mV en un determinado lugar de la dendrita, la despolarización se irá haciendo menos intensa según nos alejamos de él: –64mV, –63mV, –62mV...
Pero, ¿por qué hemos hecho referencia al cono axónico? Imaginemos por un momento que estamos situados en este lugar de la anatomía de la neurona donde están llegando, a la vez y sucesivamente, numerosos (miles, incluso decenas de miles) PEPs y PIPs. Si la suma de los diferentes potenciales postsinápticos que llegan al cono axónico en un determinado momento llega a despolarizar la membrana hasta un nivel que se denomina umbral de excitación, se producirá un potencial de acción que se propagará tal y como hemos descrito anteriormente (en los artículos precedentes de este blog).
Con el fin de simplificar las cosas vamos a utilizar sólo tres potenciales postsinápticos. Supongamos que llegan a la vez dos PEPs, uno que despolariza la neurona hasta –68mV y otro hasta –66 mV y un PIP que hiperpolariza la neurona hasta –73 mV; el resultado será, desde un potencial de membrana de –70 mV, subir 2 mV, subir 4 mV y bajar 3 mV, que globalmente supondrá una despolarización de 3 mV (–67 mV). Dado que no se ha conseguido el umbral de excitación de esa neurona, los tres potenciales postsinápticos no generarán un potencial de acción. Este hecho por el cual se integran las señales que llegan a la vez en un determinado momento se denomina sumación espacial.
Supongamos que nos encontramos en el cono axónico del ejemplo precedente. El potencial de membrana está en –67 mV, después caerá hasta –68 mV y, en ese momento, llega un PEP que despolariza la neurona hasta –66 mV. Pues bien, se producirá una integración de las dos despolarizaciones, la residual y la nueva. El resultado será una despolarización de la neurona de 4 mV por encima del nivel que se encuentra cuando llega este PEP. En consecuencia, la neurona en el cono axónico pasará a –64 mV, con lo que habrá traspasado el umbral de excitación y, por tanto, se generará un potencial de acción y, consecuentemente, un impulso nervioso. Esta integración de las señales neuronales producidas en momentos diferentes se denomina sumación temporal.
19 de septiembre de 2010
Los potenciales han saltado
Ya se ha comentado que cuando se unen los neurotransmisores a los receptores de la neurona postsináptica se produce la entrada de iones. Pero, ¿cómo sucede?
En un receptor ionotrópico, el canal que está relacionado con él (recuerde que es una proteína) se abre o se cierra, con lo que se altera el paso de iones a través de la neurona. Así, puede suceder que se abra un canal de Cl-, o que se cierre uno de K+.
Si embargo, estos receptores no son los más frecuentes, los que predominan en las neuronas son los receptores metabotrópicos. Están unidos a una proteína especial que se denomina proteína G. De esta forma, cuando se une un neurotransmisor a uno de estos receptores se produce la separación de una parte de la proteína G, la cual es capaz de unirse a un canal iónico (que se abrirá o cerrará) o, según la neurona, producirá una serie de reacciones químicas que terminarán en la formación de una sustancia, denominada genéricamente segundo mensajero, que podrá influir en el metabolismo de la célula de formas muy diferentes.

En cualquier caso, la apertura o cierre de los canales determinará una entrada o salida de ciertos iones. De esta manera podrán ocurrir varias cosas:
a) Es posible que, por ejemplo, entre Na+. En este caso se producirá en la neurona postsináptica una despolarización "que no es un potencial de acción". El potencial de membrana puede pasar de –70 a –65 mV, por ejemplo. Este potencial que se ha creado se denomina potencial excitador postsináptico (PEP).
b) También es posible que el neurotransmisor abra, por ejemplo, un canal de Cl-. Como este ión está más concentrado en el exterior que en el interior de la neurona, entrará en la misma y hará que se pase de un valor de potencial de reposo de –70 a –75mV, por ejemplo. Este potencial se denomina potencial inhibidor postsináptico (PIP).
9 de septiembre de 2010
El nuevo impulso nervioso
Los potenciales postsinápticos no responden a lo indicado en el caso de los potenciales de acción. Esto es, no se rigen por la "ley del todo o nada", sino que las amplitudes (de excitación o de inhibición) dependerán de las intensidades que las provoquen, es decir, son potenciales graduados.
Por otra parte, esta respuesta, que se inicia allí donde termine un botón terminal (sea dendrita o cuerpo celular generalmente) va decayendo progresivamente según viaja en dirección al cono axónico, la estructura cónica que se encuentra en la unión del cuerpo celular y del axón. Así, si el neurotransmisor ha generado un PIP de –75mV, según viaja y se aleja del punto inicial, esta onda de hiperpolarización irá atenuándose a –74mV, –73mV... De la misma forma, si se produce un PEP de –65mV en un determinado lugar de la dendrita, la despolarización se irá haciendo menos intensa según nos alejamos de él: –64mV, –63mV, –62mV...
Pero, ¿por qué hemos hecho referencia al cono axónico? Imaginemos por un momento que estamos situados en este lugar de la anatomía de la neurona donde están llegando, a la vez y sucesivamente, numerosos (miles, incluso decenas de miles) PEPs y PIPs. Si la suma de los diferentes potenciales postsinápticos que llegan al cono axónico en un determinado momento llega a despolarizar la membrana hasta un nivel que se denomina umbral de excitación, se producirá un potencial de acción que se propagará tal y como hemos descrito anteriormente (en los artículos precedentes de este blog).
Con el fin de simplificar las cosas vamos a utilizar sólo tres potenciales postsinápticos. Supongamos que llegan a la vez dos PEPs, uno que despolariza la neurona hasta –68mV y otro hasta –66 mV y un PIP que hiperpolariza la neurona hasta –73 mV; el resultado será, desde un potencial de membrana de –70 mV, subir 2 mV, subir 4 mV y bajar 3 mV, que globalmente supondrá una despolarización de 3 mV (–67 mV). Dado que no se ha conseguido el umbral de excitación de esa neurona, los tres potenciales postsinápticos no generarán un potencial de acción. Este hecho por el cual se integran las señales que llegan a la vez en un determinado momento se denomina sumación espacial.
Supongamos que nos encontramos en el cono axónico del ejemplo precedente. El potencial de membrana está en –67 mV, después caerá hasta –68 mV y, en ese momento, llega un PEP que despolariza la neurona hasta –66 mV. Pues bien, se producirá una integración de las dos despolarizaciones, la residual y la nueva. El resultado será una despolarización de la neurona de 4 mV por encima del nivel que se encuentra cuando llega este PEP. En consecuencia, la neurona en el cono axónico pasará a –64 mV, con lo que habrá traspasado el umbral de excitación y, por tanto, se generará un potencial de acción y, consecuentemente, un impulso nervioso. Esta integración de las señales neuronales producidas en momentos diferentes se denomina sumación temporal.
30 de agosto de 2010
El “salto” del impulso nervioso
Las señales viajan de una neurona a otra a través de una zona especial llamada sinapsis. Así, cuando el potencial de acción llega al final del axón, a lo que se denominan botones terminales, provoca la liberación de unas sustancias químicas, llamadas genéricamente neurotransmisores, que transportan las señales entre las neuronas.
En estas sinapsis los neurotransmisores se encuentran en el interior de unas vesículas que se localizan en los botones presinápticos (o terminales). La membrana de estos botones se llama membrana presináptica y esta célula nerviosa es la neurona presináptica. Por su parte, la neurona adonde va a pasar la información se denomina neurona postsináptica y sus membranas se llaman membranas postsinápticas. El espacio que separa físicamente las dos neuronas se denomina espacio o hendidura sináptica.
Estas sinapsis se denominan sinapsis químicas, para diferenciarlas de las sinapsis eléctricas en las que la unión entre las neuronas es tan estrecha que no necesitan neurotransmisor. Estas últimas son menos frecuentes que las químicas. Veamos cómo funcionan las sinapsis químicas.
Cuando el potencial de acción llega al final de la neurona, a un botón terminal, se abren unos canales de los que todavía no tenemos noticia: los canales de Ca++. Como este ión está más concentrado, en esta zona de la neurona, fuera que dentro, pasa al interior a través de los canales (1). Por su parte, la entrada de Ca++ hace que las vesículas sinápticas se muevan hacia la membrana presináptica, con la que se fusionan (2). Después de esta unión íntima el contenido de las vesículas sale a la hendidura sináptica (3). Los neurotransmisores alcanzan los receptores postsinápticos, localizados en la neurona de la membrana postsináptica, y se unen a ellos (4). Como resultado de esta unión se produce la entrada de iones cuyo resultado veremos más tarde. La transmisión finaliza cuando el neurotransmisor es degradado por unas enzimas especializadas localizadas en la hendidura sináptica; también es posible que el neurotransmisor sea recaptado (el fenómeno se denomina recaptación) por una proteína transportadora (5), ubicada en la membrana presináptica, que lo lleve al interior de la neurona que lo ha liberado y donde más tarde se incorporará de nuevo a las vesículas.

Es importante tener en cuenta que los receptores postsinápticos son específicos, es decir, un determinado neurotransmisor encajará sólo con su receptor. Así, el neurotransmisor acetilcolina podrá acoplarse a sus receptores y el neurotransmisor dopamina lo hará con los suyos, pero no podrá unirse a los receptores de la acetilcolina.
18 de agosto de 2010
El impulso nervioso
Los potenciales de acción se desplazan a lo largo del axón sin disminuir su intensidad, es decir, no se atenúan en su desplazamiento.
La transmisión del potencial de acción no es la de una corriente eléctrica: es mucho más lenta que ella y la intensidad del impulso se mantiene constante. Esto quiere decir que, en cualquier punto del axón, o hay un determinado potencial de acción, como el descrito en el anterior artículo de este blog, o no lo hay, es decir, no ocurren potenciales de acción de diferentes intensidades. Por ello decimos que siguen el principio del todo o nada.
Una vez que se ha producido un potencial de acción en un punto del axón, como consecuencia de la entrada y salida sucesivas de los iones sodio y potasio respectivamente, las corrientes de Na+ se mueven por el interior del axón provocando una apertura de los canales de sodio adyacentes y un nuevo potencial de acción.

En la figura adjunta, en A se está produciendo la despolarización a la izquierda del axón como consecuencia de la entrada de Na+, la cual se observa completa en B. Estos iones se mueven hacia zonas adyacentes y producen la apertura de los canales de Na+ de esa zona y la consiguiente despolarización. Mientras tanto, en la zona despolarizada anteriormente se regenera la situación inicial merced a la salida de K+ (lo que está representado en C).
En relación con la conducción del impulso nervioso, los axones de las neuronas pueden estar recubiertos de unas células de glía que pueden contener una sustancia lipídica denominada mielina. Los espacios sin mielina del axón se llaman nódulos de Ranvier. Pues bien, la mielina hace que los potenciales de acción sólo se generen en los nódulos, o lo que es igual, que el impulso nervioso discurra a "saltos" desde un nódulo de Ranvier a otro. Por esta razón, la propagación del impulso nervioso descrita se denomina propagación saltatoria. Si en el axón de la neurona no hay mielina la propagación del impulso nervioso se realiza de manera continua.
El hecho de tener o no mielina tiene una gran importancia en cuanto a la velocidad de transmisión del impulso nervioso. En efecto, los axones con mielina transmiten el impulso nervioso a velocidades de hasta 100 m/s, mientras que las neuronas sin mielinizar conducen los potenciales de acción a velocidades 100 veces menores y es que si hay mielina hay menos puntos que despolarizar.
Por otra parte, las neuronas con axones de gran diámetro conducen los impulsos nerviosos a más velocidad que las de axones de pequeño calibre

8 de agosto de 2010
El potencial de acción
En determinadas circunstancias es posible que se altere la permeabilidad de la membrana para el Na+. Si esto sucede es porque se abren completamente esos canales (Véase el artículo precedente en este blog) y entran masivamente estos iones, muchos más de los que penetran en reposo. Como consecuencia disminuyen las cargas positivas en el exterior y aumentan dentro, de forma que el potencial de la membrana en ese punto se va haciendo progresivamente más positivo (o menos negativo).
Este cambio del potencial de la membrana provoca, rápidamente, la apertura de los canales de K+ y estos iones empiezan a salir con más intensidad que en el reposo por razones de diferencia de concentración. Es decir, durante esta primera fase entra Na+ y sale K+, pero el número de los Na+ que entran es mayor que el de los K+ que salen. Después de, aproximadamente, 1 milisegundo (ms) se cierran los canales de sodio. En este momento la diferencia de potencial en la membrana es de unos +50 mV. Decimos que en ese punto la membrana se ha despolarizado.

Todos estos cambios que suceden en la membrana afectan solamente a los iones que se encuentran muy próximos de la misma y, por tanto, el número de iones que fluye a través de la membrana es muy pequeño. Esto supone que el equilibrio y la desigual distribución de cargas de la que partíamos se mantiene prácticamente constante. Además, el movimiento de los iones a través de la membrana y el funcionamiento de la bomba de Na+-K+ restablece las condiciones originales.
Todo este proceso que acabamos de ver en un punto del axón, en el que se ha observado un cambio en la diferencia de potencial de reposo, que sube desde –70 a 50 mV, para después descender de nuevo hasta los –70 mV, se llama potencial de acción. Por último, este potencial de acción se desplazará por el axón: es el impulso nervioso.
31 de julio de 2010
El potencial de membrana de la neurona
Cuando una neurona no funciona, está inactiva, hay en ella una pequeña diferencia de potencial entre el interior y el exterior que es perfectamente medible. Pues bien, cuando en una neurona, en la que no está ocurriendo nada, se coloca un microelectrodo en el interior del axón y se conecta a un osciloscopio se observa, después de la amplificación de la señal, que hay una diferencia de potencial entre el interior y el exterior de la neurona de unos –70 mV (milivoltios) que se llama potencial de membrana en reposo o potencial de reposo. No crea el lector que esto es exclusivo de las neuronas, al contrario, también se manifiesta en las células musculares, en las sanguíneas y en muchas otras.
Pero, ¿por qué existe esta diferencia? A ambos lados de la membrana neuronal hay una distribución desigual de cargas eléctricas que es la responsable de la diferencia de potencial: hay una mayor concentración de cargas positivas en el exterior y una mayor concentración de cargas negativas en el interior. Pero, ¿cuáles son estas cargas eléctricas?
Los iones potasio (K+) se encuentran en mayor concentración en el interior, mientras que los iones sodio y cloro (Na+ y Cl-) están más concentrados en el exterior. Por último, hay una mayor concentración en el interior de grandes aniones orgánicos como las proteínas. Sin embargo, esta situación no es estática, las partículas cargadas están en continuo movimiento y entran y salen de la célula. Esto sucede porque en la membrana de la neurona hay unas proteínas especiales que forman unos canales iónicos que permiten el paso de los iones, pero no de las proteínas porque, sencillamente, no caben.
Ahora bien, no todos los iones atraviesan la membrana con la misma facilidad, es decir, hay diferencias de permeabilidad a los iones: los K+ pasan más fácilmente (unas 30 a 40 veces más fácilmente) que los Na+. Por su parte los Cl- pasan a través de la membrana con menos problemas que los K+ y más difícilmente que los Na+. Ya hemos dicho que las proteínas no son capaces de atravesar la membrana debido a su excesivo tamaño.

Pero estos iones tienden a desplazarse según las leyes de la naturaleza, esto es, desde donde están más concentrados a donde están menos y según el hecho eléctrico de que cargas del mismo signo se repelen y de diferente signo se atraen.
*El K+ está más concentrado en el interior que en el exterior, por lo que tiende a salir como consecuencia de la diferencia de concentración, es decir, a favor de gradiente de concentración. Pero este ión, por otro lado, tiende a entrar y no salir, porque el exterior es positivo. De entre estas dos fuerzas opuestas "puede más" la primera y, en consecuencia, sale una importante cantidad de este ión.
*El Na+ tiende a entrar debido a la diferencia de concentración y también por razones eléctricas. Sin embargo, no entra una gran cantidad de iones porque la membrana no es tan permeable a estos iones como a los K+.
*El Cl- entra por razones de diferencia de concentración y tiende a salir por motivos eléctricos. Estás dos fuerzas opuestas tienen la misma intensidad por lo que, en reposo, el movimiento de estos iones está compensado.
Como acabamos de decir, en reposo (recuerde que todo esto sucede en una situación en la que no hay "actividad" neuronal) los Na+ prácticamente no atraviesan la membrana pero... siempre hay alguno que pasa y cuando esto sucede se rompe el equilibrio. Y este equilibrio hay que mantenerlo porque es el fundamento de la excitabilidad neuronal. Entonces, ¿cómo se mantiene en reposo esa desigual distribución de cargas?
A lo largo de las membranas de las células nerviosas hay unas moléculas proteicas que, trabajando permanentemente, son capaces de transportar hacia afuera el Na+ que ha entrado en la neurona y, de manera simultánea, el K+ que ha salido lo meten de nuevo. Este trabajo lo realizan a un ritmo de 3 Na+ por cada 2 K+. Estas proteínas realizan su actividad consumiendo energía (en forma de ATP) y por ello se denominan bombas. La que acabo de explicar es el funcionamiento de la bomba de sodio-potasio.
20 de julio de 2010
Algunos beneficios del estrés
Aunque el origen del estrés es diferente, los efectos siempre son similares y tienen relación con lo que se ha dado en llamar comportamiento de lucha-fuga, es decir, aumento de la glucosa sanguínea, incremento de riego sanguíneo hacia unas zonas —cerebro, músculos voluntarios y corazón— y disminución del flujo hacia otras vísceras (como las del aparato digestivo), reducción de las percepciones dolorosas, etc.
Los agentes responsables del estrés, internos o externos, alteran el equilibrio del organismo y en esto reside el peligro; consecuentemente, la gravedad del estrés dependerá de la capacidad del organismo para restablecer el equilibrio inicial necesario para mantener la vida. Si un estímulo provoca que el ritmo cardíaco suba a 130 pulsaciones por minuto, esto puede ser bueno si uno tiene que salir corriendo, pero será malo si este trabajo del corazón se mantiene varios días.
Si una persona engorda, el aumento de peso provocado por los depósitos de grasa está actuando como un agente estresante en la medida que el corazón, el sistema circulatorio, los músculos, los pulmones, etc., deberán responder ahora para desplazar “esos kilos de más”. Si usted se ha escandalizado con la medida de su báscula, puede hacer ejercicio físico, se lo agradecerán su corazón y su sistema circulatorio, pero si se pasa y frecuentemente realiza ejercicio hasta la extenuación, es probable que los efectos perjudiciales de una situación tan estresante acaben lesionando muchos órganos internos si su organismo no se recupera rápidamente del estrés.
Hay una circunstancia como el envejecimiento que tiene una relación muy estrecha con el estrés, ya que reduce la resistencia del organismo a los cambios fisiológicos producidos en esas situaciones. Sin embargo, también se puede considerar una regla general que las personas se diferencian considerablemente en su capacidad para soportar el estrés y, además, un individuo puede tolerar perfectamente una situación de estrés y aguantar muy mal otros estímulos estresantes.
En los años cincuenta del siglo XX, Hans Selye describió la situación de estrés orientándola en dos direcciones: por un lado se producirán unos cambios en el animal que le facilitarán una conducta con la que responder al estímulo desencadenante del estrés, y por otro, si las alteraciones se mantienen durante mucho tiempo, el estrés se convertirá en enfermedad.
Contrariamente a lo que se dice, además de que el estrés es necesario porque prepara al organismo para enfrentarse a una situación adversa, puede ser beneficioso por razones menos conocidas. Desde los años 60 del siglo XX muchos investigadores han puesto de manifiesto que las ratas sometidas a situaciones de estrés —producidas por descargas eléctricas— en sus primeras etapas del desarrollo, durante la vida posterior abren antes los ojos, son más precoces a la hora de adquirir coordinación motora y, en la etapa adulta, se muestran mucho más activas que las que no sufrieron estrés.
10 de julio de 2010
Los agentes del estrés
En 1939, Hans Selye, se encuentra investigando los efectos que tienen unas sustancias químicas en las ratas. Para realizar este trabajo elige dos grupos de roedores: a uno le inyecta diariamente el extracto químico, al otro grupo (control) hace lo propio con una inofensiva solución salina. Cuando comprueba los efectos, ve que el primer grupo tiene unas alteraciones muy importantes, entre las que destacan algunas úlceras pépticas y un considerable aumento del tamaño de las glándulas suprarrenales. Cuando estudia el segundo, observa, para su sorpresa, que les pasa lo mismo. ¿Qué había hecho igual en los dos grupos de ratas? La respuesta era clara: ambos grupos de animales se habían sometido al desagradable hecho de recibir un pinchazo diario, por lo que Selye investigó si las anomalías que había detectado se debían al hecho de haber soportado una situación fastidiosa.
Este científico comprobó sus hipótesis sometiendo a otros roedores a condiciones enojosas: intensos ruidos, ambientes muy fríos, sustancias tóxicas, etc. y vio que, en todos los casos, encontraba los mismos efectos que los producidos por las inyecciones. Concluyó que esas alteraciones eran la respuesta del organismo a una agresión y, usando un término de los ingenieros, habló de que las ratas sufrían “tensiones”.
No es extraño que nos encontremos ante situaciones que nos pueden causar daño o amenaza. Ante ellas se va a producir una serie de cambios en el funcionamiento general del organismo que llamamos respuesta de estrés o, más frecuentemente, estrés. Uno se puede estresar trabajando, haciendo deporte o tomando fármacos. Lo significativo es que los cambios en la fisiología corporal, como en toda emoción, afectan al sistema nervioso autónomo, sistema endocrino y a la conducta.
No es sorprendente que un amigo nos diga que está estresado cuando, en realidad, se encuentra cansado, y es que estrés y agotamiento no son la misma cosa. En primer lugar habrá que decir que una situación de estrés es fundamental para la supervivencia de las diferentes especies animales, lo que resulta perjudicial es el estrés prolongado. Los seres humanos no pueden mantenerse indiferentes ante la posible agresión de un animal, ante el examen de una oposición que va acondicionar el futuro, ante la pérdida de un trabajo… Se hace necesario preparar al organismo para que realice una actividad que le permita reaccionar ante los estímulos adversos.
Lo peligroso es soportar una situación de estrés permanente. El estrés mantenido que tiene que aguantar un controlador aéreo, un maestro, un médico de una UVI o una persona encargada del cuidado de un familiar enfermo de Alzheimer, etc., el estrés al que están sometidas estas personas es, repito, el que puede ser perjudicial o, en el peor de los casos, letal.
Las situaciones que lo provocan, los estímulos estresantes, son de naturaleza muy diversa y, en principio, podríamos clasificarlas en tres grupos: físicos, como puede ser una larga exposición a temperaturas por debajo de los 0ºC o la fractura de un hueso; químicos, como la nicotina o las anfetaminas; y estímulos psicológicos, como es el sufrimiento y ansiedad ante una operación quirúrgica que no se presume exitosa o la tristeza como consecuencia de la pérdida de un ser querido. No obstante lo anterior, hay que indicar que una misma situación puede producir estrés en una determinada persona y dejar indiferente a otra. Esto supone que el agente determinante del estrés tiene, en algunos casos al menos, un aspecto subjetivo.
1 de julio de 2010
La velocidad de los impulsos nerviosos
Las ideas más primitivas en relación con el funcionamiento del sistema nervioso suponían que el encéfalo segregaba unos “espíritus” que, a través de los nervios llegaban a los músculos y los movían. Así elucubraba, por ejemplo, René Descartes (1596-1650). No obstante, Luigi Galvani, al finalizar el siglo XVIII, demostró que los músculos de la rana podían responder a los estímulos eléctricos. Ya en el siglo XIX, Carlo Matteuci primero y Herman von Helmholtz mostraron al mundo, por lo menos al científico, la naturaleza eléctrica del impulso nervioso y que los impulsos nerviosos no viajaban a velocidades tan enormes que no se pudieran medir.
Cuando una neurona no funciona, está inactiva, hay en ella una pequeña diferencia de potencial entre el interior y el exterior que es perfectamente medible. Pues bien, cuando en una neurona, en la que no está ocurriendo nada, se coloca un electrodo en el interior del axón se observa, después de la amplificación de la señal, que hay una diferencia de potencial de –70 mV (milivoltios), que se llama potencial de membrana. No crea el lector que esto es exclusivo de las neuronas, al contrario, también se manifiesta en las células musculares, en las sanguíneas y en muchas otras.
Hay momentos en los que, en un punto del axón, se observa un cambio en esta diferencia de potencial, que sube desde –70 a 40 mV, para después descender de nuevo hasta los – 70 mV. Este cambio es el llamado potencial de acción o impulso nervioso, y es debido, respectivamente, a la entrada y salida sucesivas de iones cargados positivamente (Na+ y K+) Este cambio va discurriendo por todos y cada uno de los puntos del axón hasta llegar al final de la neurona: es el impulso nervioso discurriendo por el axón de la neurona.
No es infrecuente oír por la televisión, incluso en programas de ¿divulgación científica?, que los impulsos nerviosos viajan a la velocidad de la luz. Es claro que este asunto es extraordinariamente atrayente y que fue objeto de elucubración primero y de estudios serios después, y desde hace mucho tiempo, por los científicos. En el siglo XVIII, Albrecht von Haller consideró que la velocidad del impulso nervioso en las neuronas humanas era de unos 50 m/s (metros por segundo); en el siglo XIX, Johannes Müller estimaba que debían ir a gran velocidad porque “el pensamiento es muy rápido”. En el mismo siglo, el ya citado Helmholtz valoró la velocidad del impulso nervioso en 26 metros por segundo.
Hoy sabemos que el impulso nervioso se propaga a una velocidad que, dependiendo de las neuronas, oscila entre los 5 y los 120 m/s, lo que está bastante lejos de la velocidad de la luz que es de 300000 km/s. En la especie humana podemos considerar que, por término medio, la velocidad de conducción del impulso nervioso, en las neuronas que contactan con las células musculares, es de unos 60 m/s. Es claro que hay una pequeña diferencia entre la realidad científica y las informaciones (¿?) televisivas y, es evidente que muchos de los científicos del siglo XIX sabían más de esto que algunos divulgadores del siglo XXI.
22 de junio de 2010
La esquizofrenia y la genética
Aunque parezca raro, el 1% de la población mundial padece un conjunto de síntomas que, en mayor o menor medida, se manifiestan con signos como poseer ideas tan delirantes como la de estar sometido a ilusiones persecutorias, padecer alucinaciones auditivas que dirigen a la persona en una determinada dirección, tener pensamientos eventualmente incoherentes, mostrar incapacidad para reaccionar emocionalmente como lo haría la mayor parte de la población, presentar un comportamiento incomprensible, etc. Es la esquizofrenia, término que fue adoptado en los primeros años del siglo XX para referirse a un trastorno cuyo síntoma más significativo era la ruptura de la integración entre pensamiento, emoción y acción, y es que esquizofrenia quiere decir separación de funciones psíquicas.
Las discusiones sobre la importancia de la naturaleza y del ambiente se han centrado durante mucho tiempo en esta enfermedad maldita. El psicoanálisis, en un primer momento, no trataba a estos enfermos y el padre de esta disciplina, Freud, se atrevió a conjeturar que la esquizofrenia era la consecuencia de la represión ejercida sobre los impulsos homosexuales. ¡Toma ya!
La proporción de esquizofrénicos indicada antes es igual en todos los estudios realizados y esto es muy sorprendente. Cabe pensar que siendo un fenotipo debido a muchos genes, los alelos se expresen de manera favorable, positiva, excelente, siempre y cuando no estén muchos de ellos juntos en una misma persona; en ese caso, formarán una mezcla explosiva. Quizá esto explique que tener algunos de estos alelos sea bueno y, por tanto, que la evolución los haya mantenido desde hace cientos de miles de años. Por otra parte, esta hipótesis es concordante con un hecho tan interesante como que personajes tan geniales como Immanuel Kant e Isaac Newton presentaron una versión atenuada de la enfermedad.
Los fríos datos estadísticos nos dicen que esta conducta tiene un fuerte componente familiar. Si usted no conoce ningún miembro esquizofrénico de su familia, la probabilidad de que en algún momento de su vida se le manifieste la patología es del 1%, pero si alguno de sus parientes la padece, el riesgo de que usted la sufra aumenta considerablemente.
En el último tercio del siglo XX se hicieron numerosos “estudios de familias” que afectaron a varios miles de esquizofrénicos. Las estimaciones indicaron claramente que si una persona padece la enfermedad, la probabilidad de que alguno de sus hijos sea esquizofrénico es del 13%. Sin embargo, si los dos progenitores padecen la enfermedad, el riesgo de que uno de sus hijos la manifieste asciende hasta el 46%.
En estos trabajos se suele utilizar el valor de la concordancia; así, en una muestra de gemelos en la que al menos uno está afectado, la concordancia hace referencia al porcentaje de parejas que coinciden en la enfermedad. Por ejemplo,
Pues bien, estudios recientes realizados en Europa y Japón indican que la concordancia de la esquizofrenia entre gemelos idénticos es de alrededor del 50% (si en 100 parejas en las que al menos uno de los miembros es esquizofrénico hay 50 en las que ambos miembros presentan la enfermedad, la concordancia es del 50% ) y en el caso de los mellizos es del orden del 15%. En cualquier caso, estos estudios suponen una gran influencia genética, pero también implican que, en el caso de los gemelos monocigóticos, en el 50% de los casos no hay concordancia para la enfermedad. Esto supone que factores no genéticos son fundamentales para que se desarrolle la patología.
¿Y si le seguimos la pista a una pareja de gemelos discordantes para la esquizofrenia? Los hijos del gemelo enfermo tienen una gran probabilidad de ser esquizofrénicos como su progenitor, pero los del gemelo no afectado presentan el mismo riesgo, a fin de cuentas tiene unos “genes para padecer la esquizofrenia” que no ha manifestado. De esto se colige que tener los genes para sufrir una enfermedad como la que tratamos es absolutamente necesario pero, afortunadamente, no es suficiente.
Desde siempre se han buscado influencias ambientales a la hora de explicar por qué surge la esquizofrenia. Hace mucho tiempo se sabe que factores no genéticos guardan relación con la enfermedad: padecer la gripe durante el embarazo, la no ingestión de ácidos grasos esenciales, la disminución del aporte de oxígeno a causa de una preeclampsia aumenta hasta nueve veces el riesgo de ser esquizofrénico, etc.
13 de junio de 2010
El cerebro del músico
Los músicos son un modelo excelente a la hora se investigar la importancia que tiene la experiencia en el modelado cerebral. Es por eso que estos artistas constituyen unas personas excepcionales en los estudios que tienen que ver con la plasticidad cerebral.
Las técnicas de neuroimagen cerebral permiten distinguir las diferencias anatómicas y fisiológicas de los cerebros de diferentes individuos o de la misma persona en situaciones distintas. Esto hace posible, por ejemplo, comparar los cerebros de una persona leyendo o con los ojos cerrados o, en relación con el título de este artículo, de un virtuoso del piano y el de un analfabeto musical, y cotejar la actividad neural del intérprete cuando está relajado y cuando interpreta una pieza de Sergéi Rajmáninov.
Schlaug, Gottfried y sus colaboradores comprobaron, a mediados de la década de los noventa del siglo XX, una diferencia anatómica fundamental entre los músicos y los no músicos: la masa nerviosa anterior y media que conecta los dos hemisferios cerebrales, el denominado cuerpo calloso, era mayor en los primeros. Esta diferencia morfológica implica, de alguna forma, otra de carácter fisiológico ya que supone una, probablemente, mayor comunicación entre los dos hemisferios cerebrales en los músicos. Lo que, por otra parte, no debe sorprendernos en la medida que en los cerebros de músicos es mayor la sustancia gris de las cortezas motora y auditiva y del cerebelo. Dicho de otra forma, utilizando la comparación es fácil identificar el cerebro de un músico.
Ahora bien, cabe preguntarse: ¿los cambios anatómicos referidos son una consecuencia de la actividad musical o, por el contrario, las personas con esas modificaciones en la morfología encefálica son las que muestran habilidades musicales?
Los científicos citados antes demostraron que los cambios cerebrales a los que me he referido eran tanto más significativos cuanto antes se hubiera comenzado el aprendizaje musical y cuanto más tiempo se dedicara a la práctica musical.
Y es que no es raro que la plasticidad cerebral, propia de muchas regiones cerebrales, explique que los ejercicios ante un piano o una guitarra, por ejemplo, supongan un mayor desarrollo de la corteza motora.
2 de junio de 2010
Inteligencia y genes
Todo el mundo ha oído hablar de los tests de inteligencia. El CI (cociente de inteligencia o capacidad intelectual, como prefiera) expresa la relación entre la edad mental y la cronológica. Muchos han dicho que esos tests no sirven para nada, que no discriminan lo suficiente, que no... Es evidente que no evalúan todas las capacidades de la inteligencia, tienden a medir la capacidad de pensar, no el valor de una persona. Los tests de inteligencia no fueron creados a partir de una supuesta teoría de la inteligencia y, por esto, aunque clásica, es muy acertada la definición que de la inteligencia dio, en 1923, el psicólogo E.G. Boring: “La inteligencia es lo que miden los tests de inteligencia”. No obstante, sirven y sólo sirven para lo que fueron diseñados: prever la capacidad de los niños para avanzar en su educación, o si lo prefiere, y quizá exagerando un poco, los test sobre el CI predicen bastante bien las calificaciones educativas.
Hoy día es más frecuente hablar de capacidad cognitiva general y en ésta, hay un factor g, general, que nos indica que las personas que lo tienen alto realizan muy bien los diferentes aspectos del test, y las que lo tienen bajo mal. Por otra parte, en los tests de inteligencia hay destrezas específicas que son de valoración significativa: el cálculo numérico, la fluidez verbal, la memoria, etc. Por tanto, la inteligencia es el resultado de la combinación de capacidades generales y específicas.
¿Hay algo en la anatomía de una persona que nos puede servir para descubrir si es inteligente? La respuesta es casi afirmativa porque guarda relación con el tamaño cerebral. Esto no parece extraño ya que es lógico pensar que las personas más inteligentes tengan más neuronas, o más conexiones entre ellas, que las normales. No se preocupe si conoce gente inteligente de poca cabeza o cabezotas tontos, porque la correlación entre la inteligencia y el volumen cerebral es sólo de 0,4, lo que supone que hay un amplio espacio para que cualquier persona se tranquilice, sea cual sea el tamaño de su cabeza.
En los años 2001 y 2002 se publicaron dos trabajos (de los equipos científicos del finlandés P.M. Thompson y la holandesa D. Posthuma) en los que había una correlación enorme (0,95) en el tamaño de la sustancia gris cerebral entre los gemelos idénticos y una considerablemente menor (0,50) entre mellizos. Esto explicaba los títulos de los dos artículos científicos: “Influencia genética en la estructura del cerebro” y el más concluyente de Posthuma: “La asociación entre el volumen cerebral y la inteligencia es de origen genético”.
Si analizamos estadísticamente la correlación en el cociente de inteligencia entre parejas de personas, los resultados son bastante concluyentes: si no están emparentados es de 0, si lo están en tercer grado es 0,15, si en segundo grado 0,30 y en primer grado 0,45. Finalmente, la correlación entre mellizos es 0,60 y entre gemelos idénticos 0,85. Tenemos, por tanto, que cuanto más cercanía genética, el CI es más parecido. Aunque hay numerosas influencias ambientales que pueden afectar a la inteligencia, parece evidente que este fenotipo es familiar.
Se han realizado numerosos trabajos para discernir la influencia por separado de los genes y el ambiente. Así, al estudiar 58 parejas de gemelos monocigóticos criados en ambientes distintos (porque han sido dados en adopción) se ha demostrado una muy alta correlación en el CI (0,75).
Por otra parte, en un estudio (Colorado Adoption Proyect) publicado en 1997, dirigido por el gran genetista de la conducta Robert Plomin, se demostró que las correlaciones en el CI entre padres y sus hijos biológicos (grupo control) eran de 0,20 en los primeros años de la niñez, se mantenía este valor en la infancia media y ascendía a 0,30 en la adolescencia; era una situación en la que se compartían genes y ambiente. El mismo patrón de correlación en el CI se daba entre las madres biológicas y sus hijos dados en adopción; en este caso sólo se compartían los genes, el ambiente era, obviamente, distinto. Finalmente, estos estudios también demuestran que las correlaciones entre los hijos adoptivos y sus padres adoptivos (sólo tienen en común el ambiente) son cercanas a cero, lo que expresa que el ambiente compartido no tiene una influencia significativa en el cociente de inteligencia. Todo ello implica que la semejanza entre padres e hijos en el CI es debida a los genes.
23 de mayo de 2010
El pez cebra en la investigación neurocientífica
Estoy harto de escuchar en las tertulias radiofónicas a reputados periodistas que se burlan de que un determinado grupo de científicos está investigando “no sé qué” en cierto insecto o en una rata. Se ríen de lo “interesante” del tema en cuestión. En realidad, ellos, como muchos miembros de nuestra sociedad, políticos a la cabeza, son unos perfectos ignorantes de que muchos de los grandes avances científicos se han realizado, se están haciendo y se practicarán en animales, donde se estudiarán fenómenos que ellos, ni con un gran esfuerzo de imaginación, serían capaces de sospechar. Pondré dos ejemplos sencillos .
Importantísimos estudios sobre el origen, transmisión y propagación del impulso nervioso (algo que parece importante) se realizaron en el axon gigante del… calamar. Por otro lado hay que decir que una de las formas más sencillas y universales de aprendizaje en los seres vivos es la habituación, aprendizaje que consiste en una reducción en la amplitud de la respuesta que sigue a la repetición de un determinado estímulo. Pues bien, el animal modelo en estos estudios es otro molusco: pertenece al género Aplysia.
Pues resulta que, en este contexto, hay una especie que está siendo utilizada por los neurobiólogos desde hace pocos años, una especie que conocen todos los aficionados a la acuariofilia, el pez cebra, al que los científicos denominan Danio rerio. Empezó a utilizarse en la investigación biológica al iniciarse la década de los setenta del siglo XX y fueron las investigaciones genéticas de George Streisinger, de la Universidad de Oregón, las que comenzaron a poner de moda este pececito.
¿Qué tiene de particular este animal? El pez cebra es, obviamente, un vertebrado y esto, desde el punto de vista científico, supone un “salto” cualitativo muy importante en cuanto a la complejidad de la especie: no es lo mismo hacer investigaciones en invertebrados que en peces, reptiles, mamíferos, primates.... y, consecuentemente, hay un mayor parecido de muchas estructuras de este animal con las humanas. Por otra parte, los individuos de esta especie tienen una gran capacidad reproductora, lo que permite obtener muchas generaciones en poco tiempo. Además, desde los primeros estados del desarrollo, el pez cebra es transparente, lo que facilita el seguimiento de algunas alteraciones orgánicas. A esto habría que añadir que en 24 horas el embrión pasa del estado de dos células hasta un estado larvario donde se pueden identificar, sin ningún problema, diversas estructuras, entre ellas el ojo. Por último, es muy fácil conseguir mutantes de pez cebra: en el Instituto Max Plank de Biología Evolutiva de Tübingen hay unas 400 variedades diferentes de Danio rerio, que se han obtenido mediante tratamientos que han provocado alteraciones en su información genética. Así, hay ejemplares con defectos en el sistema nervioso, en los músculos, órganos internos, etc. Hay razas que carecen del sentido del tacto, o son peces ciegos, o tienen una cierta incapacidad para desplazarse hacia adelante, etc.
Más de uno se asombraría de que muchas de las investigaciones que se están realizando sobre la retinitis pigmentosa, una alteración de la retina que provoca su desprendimiento progresivo y que termina produciendo ceguera, se están haciendo en el pez cebra. Esta enfermedad es una de las principales causas de ceguera en las poblaciones humanas de los países industrializados. Curiosamente, esta anomalía está producida en el pez cebra, y en el hombre, de la misma forma: la muerte de las células receptoras visuales que están situadas en la retina, los bastones y los conos. Esta línea de investigación es la que en la actualidad están siguiendo los científicos Beatrix Benz y Stephan Nauhauss, en Zurich.
Muchas investigaciones sobre el síndrome Usher 1-B, que se manifiesta casi siempre en los humanos con una sordera congénita (total o parcial) se están realizando en estos peces, y es que podemos decir que, en los aspectos fundamentales, el oído de este animal funciona de manera muy similar al del hombre (los peces tienen además una estructura auditiva especial denominada línea lateral). En esta dirección está trabajando en la actualidad el equipo de la bióloga Teresa Nicolson, en la Universidad de Tübingen,
En fin, Danio rerio no sólo es un vertebrado primitivo que adorna los acuarios de todo el mundo sino que también es un espléndido material biológico con el que se realizan investigaciones por el bien de nuestra especie.
13 de mayo de 2010
La ezquizofrenia de un genio
Una mente prodigiosa es un libro, que apareció en España en 2002, en el que se relata la vida de John Forbes Nash (1928), y por el que su autora, Sylvia Nasar, fue galardonada con el National Book Critics Circle Award y finalista del Premio Pulitzer. A los que les guste el cine recordarán que en el año 2001 fue estrenada en España la película de Ron Howard titulada Una mente maravillosa; interpretada por Russell Crowe obtuvo cuatro Oscar, entre los que se encontraban dos de los más prestigiosos: a la mejor película y al mejor actor.
John Forbes Nash fue un matemático excepcional: estudios sobre teorías de juegos, sobre variedades algebraicas, sus trabajos sobre ecuaciones diferenciales con derivadas parciales, etc. El matemático William Browder ha considerado que Nash es el mayor especialista en numerología que jamás ha existido en el mundo.
A pesar del prestigio conseguido en vida, Nash no obtuvo un galardón matemático hasta 1978; fue el Premio de Teoría John von Neumann de la Sociedad de Investigación de Operaciones y el Instituto para la Ciencia de la Dirección. Después , recibió el premio Nobel de Economía.
La aparición de la esquizofrenia en el matemático fue en la década de los 50, pero lo que parece claro, con las noticias que se aportan en el libro de Sylvia Nasar, es que Nash siempre tuvo una personalidad “especial”. Desde muy pronto se manifestó como un hombre intelectualmente vanidoso y más presuntuoso y extravagante que sus compañeros universitarios, un hombre que despreciaba muchos de los conocimientos de los demás porque aprender demasiadas cosas de otros le impedía ser original.
Nash no era simpático, pero inspiraba respeto y su conducta infantil o, si se prefiere, adolescente, le valió el mote de “profesor niño” cuando tenía 23 años y era docente auxiliar del MIT. Odiaba a los psiquiatras y era despectivo con los alumnos y con sus compañeros. Llegó a decir: Hay pocos genios en el MIT: yo, por supuesto, y también Norbert Wiener. Incluso es posible que Wiener ya no lo sea, pero hay pruebas de que lo fue en el pasado. Su colega Warren Ambrose le calificó de brillante, engreído, infantil, irreflexivo e indisciplinado.
Tuvo un hijo, John David, de su relación con Eleanor Stier, con la que no se casó, ni ayudó económicamente, ni se ocupó del niño; la madre tuvo que cederlo a una serie de familias, aunque después lo recuperó. Nash parece que también tuvo algún que otro escarceo homosexual. Lo cierto es que, empero, se casó con Alicia Larde —a la que conoció mientras estaba de profesor en el MIT—, que tanto había de contribuir a la curación o mejora de su enfermedad. Con Alicia tuvo un hijo, al que también llamaron John, que manifestó la misma enfermedad que su padre y que se doctoró en matemáticas por la Universidad de Rutgers.
Sylvia Nasar saca a flote muchos planteamientos, ya científicos, ya culturales, en lo que se refiere a la enfermedad, diagnosticada en 1959 como esquizofrenia paranoica. También, en esta excelente biografía, queda perfectamente explicada una breve historia de la terapéutica médica utilizada para tratar la enfermedad del matemático, inicialmente con toracina y psicoanálisis, después, los dolorosos tratamientos con insulina —ideados en la década de los 20 por el vienés Manfred Sackel— y, finalmente, el uso de fármacos antisicóticos. Nasar nos enseña cómo la enfermedad parece remitir y vuelve a rebrotar con signos espectaculares.
Lo cierto es que mientras duró, y dura, la esquizofrenia, cuenta Fagi Levinson, todo el mundo quería ayudar a Nash: era una mente demasiado brillante para dejar que se perdiera.
4 de mayo de 2010
Adaptación y habituación
Cuando entramos en una habitación que huele de manera desagradable detectamos el olor pero, al rato, casi no lo percibimos. ¿Qué pasa con las señales nerviosas?
Esto también sucede con otros estímulos sensoriales. A veces nos ponemos una determinada prenda que en contacto con la piel nos “pica” en un primer momento y, poco después, no nos enteramos de que la llevamos puesta. O nos echamos colonia y, de nuestro alrededor, los únicos que no la olemos somos nosotros.
La solución a la pregunta es, en principio, bastante simple: o los receptores dejan de responder a los estímulos o nosotros dejamos de percibir la estimulación. Y esto no es ilógico, es decir, encaja bastante bien en los diferentes aspectos evolutivos.
Los seres vivos necesitamos tener información de lo que sucede en el exterior para hacer frente a las diferentes situaciones; lo que es innecesario es que se nos recuerde de una manera machacona. Piense, por ejemplo, en que, desde el punto de vista biológico, es más que suficiente saber si un determinado olor procede de un depredador que se acerca hacia nosotros o es el de una presa de la que vamos a dar buena cuenta. No es preciso mantener el olor permanentemente.
Así, las neuronas olfativas son receptores del olor capaces de responder a determinadas sustancias volátiles. Las respuestas son potenciales generadores que terminan en potenciales de acción cuya frecuencia es tanto mayor cuanto mayor es la concentración de la sustancia olorosa, esto es, a mayor concentración de sustancias olorosas mayor número de impulsos nerviosos viajan por las neuronas y una mayor percepción es “traducida” por nuestra corteza sensorial olfativa.
Sin embargo, el mantenimiento de los estímulos olfativos acaba produciendo una reducción progresiva de los impulsos nerviosos, es decir, las neuronas receptoras manifiestan una adaptación sensorial. Esto suele suceder porque la estimulación repetida de la neurona receptora, en este caso, produce la inactivación de los canales de Na+ que antes se abrían ante los estímulos químicos. Dicho de otra forma, la neurona deja de ser estimulada por las sustancias químicas volátiles.
En otras situaciones, muchos receptores táctiles por ejemplo, los receptores continúan permanentemente respondiendo a los estímulos, pero es el individuo es el que deja de hacerlo: es la habituación, un aprendizaje que genera una modificación del funcionamiento de la sinapsis en el sistema nervioso central o fenómenos fisiológicos mucho más complicados. Esto es, en este caso se aprende a no ser estímulado por un estímulo que se repite.
22 de abril de 2010
Factores insulínicos y cerebro
La insulina es una hormona conocida por todos y asociada a una enfermedad que denominamos diabetes; guarda, por tanto, relación con la glucemia, esto es, con el contenido de glucosa en la sangre. La insulina es la única hormona que se encarga de bajar la concentración de glucosa en la sangre y, en consecuencia, es imprescindible a la hora de mantener la glucemia.
Es menos conocido que hay neuronas que segregan insulina y otros péptidos semejantes que se denominan factores insulínicos o IGF. Aún es menos sabido que en algunos invertebrados tan sencillos como el nematodo Caenorhabditis elegans hay más de treinta de estos factores y 10 en mamíferos, lo que implica que, desde el punto de vista evolutivo, deben ser un acierto fisiológico y, como resultado, fundamentales.
Una de estas sustancias guarda relación con el desarrollo cerebral. Así, en ratas manipuladas genéticamente, la falta de uno de estos factores, denominado IGF1, produce un cerebro anormalmente pequeño mientras que su exceso genera megacerebros.
En las primeras etapas de la ontogénesis las neuronas producen el IGF1 necesario para estimular la neurogénesis, Por su parte, alcanzado un determinado nivel de desarrollo, cesa la fabricación general de este factor que se circunscribe a zonas concretas del cerebro. Bien es cierto que, entonces, las neuronas disponen de los factores en la sangre y, en el caso de los vertebrados, las principales células fabricantes de los mismos son las pancreáticas, las hepáticas y las intestinales. En nuestra especie, la producción de ese factor decrece desde los 30 años aproximadamente.
Se ha visto que en ciertas patologías que afectan al sistema nervioso, tales como la depresión, accidentes vasculares, demencia y otras, los niveles sanguíneos de IGF1 estaban alterados en relación con los normales. Pero, ¿por qué cambia la concentración de IFG1 sanguíneo en las patologías neurodegenerativas?
Hay científicos que sostienen la hipótesis de que esta sustancia es de protección neuronal, lo que implica que IGF1 podría bajar su concentración antes de la enfermedad o bien al revés: las patologías neurogenerativas provocarían la disminución del citado factor insulínico.
La mayor parte de las investigaciones señalan que los cambios neuronales patológicos son la consecuencia del déficit de IGF1 pero, fuera como fuera, la administración de esta sustancia es posible que tenga efectos terapéuticos.
Suscribirse a:
Entradas (Atom)